


Eşcinsellik Bulaşıcı mı?: Eşcinselliğin Patojen Teorisi

[Epistemik Statü: Spekülasyon]
Bu yazıda, “evrimsel bir paradoks” olan eşcinselliğin, patojen (virüs, mikrop, parazit vb.) bulaşımından kaynaklanma ihtimalinin var olan eşcinsellik teorileri arasında en güçlü adaylardan biri olduğunu savunacağım.
1. Dışlayıcı eşcinsellik, Eşcinsel (veya eş cinse yönelik) davranış ve Evrimsel Paradoks
1.1. Erkek Eşcinselliği, Kadın “Eşcinselliği”
Eşcinsellik ve eş cinse yönelik davranış ayrımına geçmeden önce, yazı boyunca, “eşcinsellik” ile “erkek eşcinselliği”ni kast edeceğimi belirtmeliyim. Bunun nedeni, kadın ve erkekler arasındaki cinsel davranış örüntüsü farkı nedeniyle “eşcinsellik” kelimesinin iki cinsiyet için aynı şeyi ifade etmemesi.
Erkekler, cinsellik konusunda daha kategori spesifikken (eşcinsel olduğunu söyleyen bir erkek karşı cins tarafından uyarılmıyor veya onunla ilişkiye girmiyorken), kadınlar kategori spesifik bir cinsel davranış/uyarılma örüntüsüne sahip değil. Örneğin, (Bailey, 2009), erkek ve kadınların cinsel tercihleri ve cinsel uyaranlara (eşcinsel cinsellik görüntüsü veya düzcinsel cinsellik görüntüsü) fizyolojik cinsel uyarılma tepkilerinin farklı olduğunu gösteriyor. Erkekler, kendi tercih ettikleri cinsiyet dışındaki uyarılmayı kadınlara göre çok daha az yaşıyor. Görülebileceği üzere, eşcinsel ve heteroseksüel kadınlar hem kadınlar hem erkekler tarafından uyarılıyor.

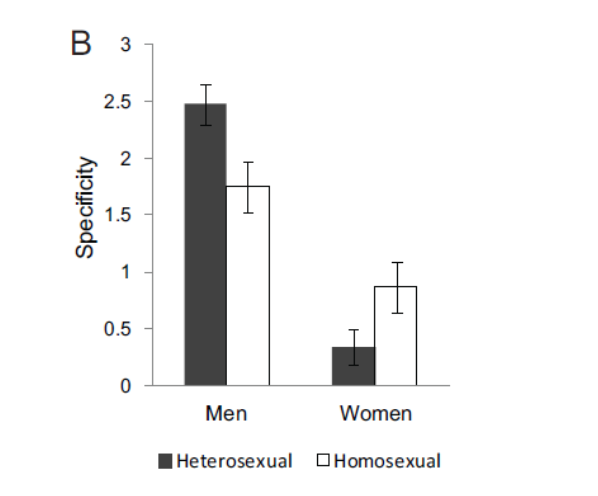
Benzer biçimde, (Sylva et al., 2014), erkek ve kadınların cinsel tercih ve uyarılma ilişkisini beyin görüntülemesi üzerinden inceliyor ve yine erkeklerin kategori spesifik uyarılma örüntüsüne sahipken kadınların çok daha düşük kategori spesifiklik gösterdiğini buluyor.
Bunun yansımasını lezbiyen cinsel davranışında da görebiliyoruz. Örneğin, (Diamant, 1999), kendini lezbiyen olarak tanımlayan yaklaşık 7000 kadın üzerinden yapılan bir ankette, en az bir tane erkek partneri olan lezbiyen oranını %77 ve vajinal ilişkiye girme oranını %70 buluyor.
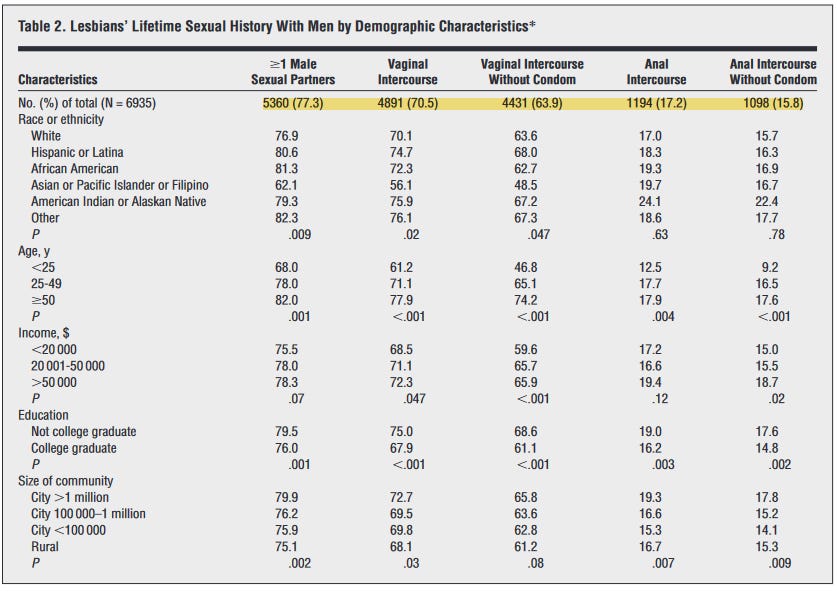
Benzer biçimde, (Bailey, 2003), lezbiyen ve biseksüel kadınların cinsel geçmişine dair çalışmasında, örneklemin %85’inin son on yılda en az bir erkekle birlikte olduğunu gösteriyor.

Bunun yanında, yakın dönemli bir genom-çapı ilişki çalışması, erkek ve kadın eş cinsel davranışının farklı etiolojileri olduğuna işaret ediyor zira kadın ve erkek eş cinsel davranışıyla ilgili SNP’ler ortak değil. (Ganna et al., 2019)
Bu nedenlerden ötürü “eşcinsellik” iki cinsiyet için aynı şeyi ifade etmiyor ve ikisini farklı şekilde açıklamak gerekiyor.
1.2. Evrimsel bir paradoks olarak eşcinsellik
Eşcinselliğin evrimsel paradoksuna gelmeden önce, genellikle yapılmayan fakat çok önemli bir ayrımı yapacağım: (Dışlayıcı) Eşcinsellik ve Eşcinsel (veya Eş Cinse Yönelik) Cinsel Davranış ayrımını.
Eşcinsel veya eş cinse yönelik cinsel davranış, adından da anlaşılabileceği üzere, belirli bir cinsiyetteki bireyin kendi cinsiyetinden bir bireyine yönelik cinsel davranışıdır. Örneğin, erkek bir aslanla ilişkiye giren bir erkek aslan, eşcinsel davranış sergiliyor demektir. Eşcinsel davranış bir eylemdir. Bunun aksine, eşcinsellik bir karakteristik özelliktir ve istikrarlı/sabittir. Dişi aslanlarla ilişkiye giren bir aslanın eşcinsel davranışta bulunması onu eşcinsel yapmaz. Fakat bir erkek aslan, dişilerle ilişkiye girmiyorsa ve sadece eşcinsel davranışta bulunuyorsa, bu onu (dışlayıcı) eşcinsel yapar. Aynı şekilde, hapse giren bir erkeğin orada gönüllü olarak erkeklerle ilişkiye girmesi onu eşcinsel yapmaz.
Bu ayrım önemli, çünkü bunlardan dışlayıcı eşcinsellik “evrimsel paradoks” içeriyor: Eşcinsellik evrimsel/genetik olamaz zira eşcinselliğin evrimsel uyum maliyeti yüksektir, birkaç nesilde silinmesi gerekir fakat eşcinsellik dünyada görece yaygın (%2-4).
Önce eşcinselliğin neden genetik/evrimsel olmaması gerektiğine dair daha açık bir açıklama yapıp, sonra bu paradoksu evrimsel olarak çözmeye yönelik girişimlerden bahsedeceğim.
Öncelikle, eşcinselliğin temelde genetik olmamasını beklememiz gerekiyor zira ikiz çalışmaları, eşcinselliğin sadece %20 kalıtsal olduğunu gösteriyor (Whitehead, 2012), eşcinsellik büyük ölçüde paylaşılmayan çevre kaynaklı gibi görünüyor.

Bundan da önemlisi, gen aktarım katsayıları uymuyor. Diyelim, kişilerin eşcinsel olmasına neden olan bir gen var ve bir toplumun yarısı bu gene sahip; bunun yanında eşcinsellik genine sahip olanların çocuk yapma oranı da eşcinsel olmayanların yarısı, yani 0.5 bir uyum maliyetine sahip. İkinci nesilde bu gene sahip kişi oranı %25’e, üçüncü nesilde %12.5’e… düşer. 10. nesilde yaklaşık yüz binde 1’e kadar geriler. Fakat bu bizim bildiğimiz %2-4 oranla epey uyumsuz.
Dahası, biz 0.5 gibi görece düşük bir uyum maliyetinden bahsettik. Fakat empirik çalışmalar bundan da düşük bir oranı işaret ediyor. Örneğin, (Apostolou, 2022), ABD’de görece yakın zamanlı iki veritabanını kullanarak cinsel yönelim ve çocuk sayısını inceliyor ve çağdaş eşcinsel erkeklerin neredeyse hiç çocuk sahibi olmadığını buluyor.

Benzer biçimde, (Vasey et al., 2014), Samoa’da (fa’afafine olarak adlandırılan) eşcinsel erkeklerin hiç çocuk yapmadığını buluyor.
Buna karşı getirilen bir itiraz, cinsel yönelimlere baskının daha yüksek olduğu zamanlarda erkeklerin evlenip çocuk yaptığı şeklinde. Bu iddia kısmen doğru. Eşcinsel erkeklere evlenip çocuk yapma baskısı mevcut olsa da, bu çocuk sayısıyla ilişkili bir baskı değil, eski zamanlarda (veya şu an cinsel tercihlerde baskıcı toplumlarda) erkeklerin evlenseler bile daha az çocuk yapmalarını bekleyebiliriz. Nitekim öyle de. Örneğin, (Moran, 1973), evli eşcinsel erkeklerin çocuk sayılarının heteroseksüellere göre çok daha düşük olduğunu buluyor ve uyum yükünü 0.2 olarak hesaplıyor.
Bazıları, eşcinselliği akraba altruizmiyle açıklamaya çalışıyor. Bireyin kendisi üremese bile eğer yakın akrabasının (i.e, kardeşi) üremesine yardımcı olursa, paylaşılan genlerden dolayı bu genetik olarak adaptif olabilir. Bu teorinin en büyük zorluğu yine niceliksel genetikle alakalı. Kişinin kendi çocuğuyla genetik yakınlığı .5’ken kardeşinin çocuğuyla genetik yakınlığı .25, yani baba-çocuklar genlerinin %50’sini paylaşırken amca-yeğenler genlerinin %25’ini paylaşıyor. Bu da, yapmadığı her çocuk için akrabalarının üreyip çocuklarının da üreme çağına gelmesine iki kat fazla katkıda bulunmasını gerektiriyor. Yani, eşcinsel bir erkeğin, en az dört yeğeninin çocuk yapacak yaşa kadar yaşamasına yardım etmesi gerekiyor ki iki çocuk yapmışa denk olsun. Fakat bunu gösteren empirik bir emare yok. (Bobrow ve Bailey, 2001)
Patojenler ve Eşcinsellik
Eşcinselliğin, virüs, bakteri, mikrop gibi bulaşıcı patojenler kaynaklı olduğu fikri ilk kez (Cochran, Cochran, Ewald, 2000)’de ortaya atılıyor. Bu, çok daha geniş kapsamlı ve önemli bir fikrin örneklemelerinden biri. Hem (Cochran, Cochran, Ewald, 2000) hem de Paul Ewald’un Plague Time: The New Germ Theory of Disease’inde ortaya atılan teori, şu anda nedenini bilmediğimiz pek çok kronik ve kronik olmayan hastalığın asıl sebebinin patojenler olduğunu ileri sürüyor. Eğer bir hastalık veya kondisyon,
Yaygınsa,
Evrimsel maliyeti varsa,
Tüm çabalara rağmen nedenini bulamadıysak,
Bunun arkasında bulaşıcı bir mikroorganizmanın olma ihtimali yüksek.
Bu “Yeni Patojen Teorisi” epey verimli bir teori. Örneğin, (Ewald, 2009)’da bazı kanserlerin patojenlerle ilişkisini görebiliyoruz; kanserlerde patojenlerin oynadığı rol gittikçe daha iyi anlaşılıyor ve Ewald göre kanserlerin yaklaşık %95’inin patojenlerle ilişkisi bulunacağını iddia ediyor.

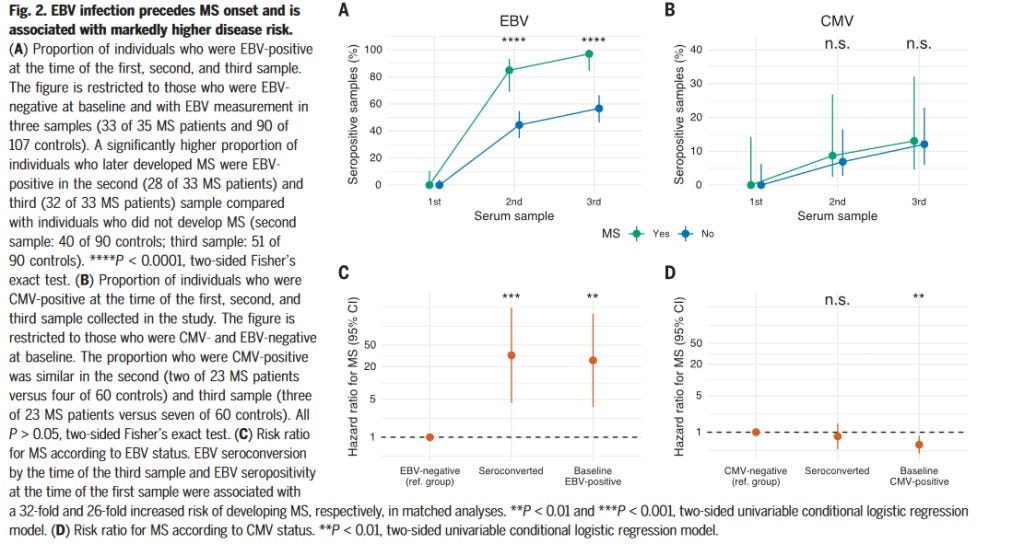
Yakın zamanlı bir örnek olarak, (Bjornevik et al., 2022), Epstein-Barr Virüsü enfeksiyonunun MS Hastalığı riskini ciddi ölçüde artırdığını buluyor.
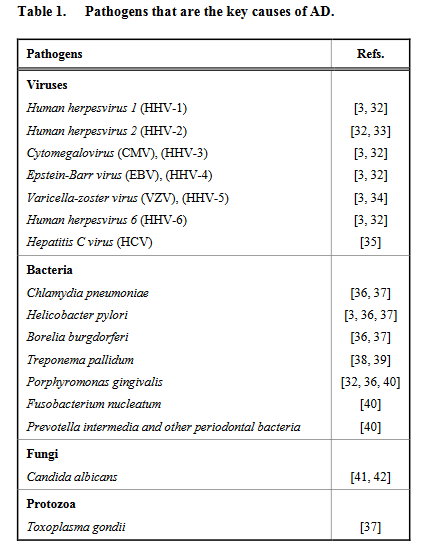
Benzer biçimde, (Balin et al., 1998), 19 Alzheimer hastasının 17’sinin kadavrasında Chlamydia pneumoniae bakterisinin izine rastladığını ve patojenlerin Alzheimer hastalığında söylemesinin ardından geçen 20 senede, c. Pneumoniae dahil pek çok patojenin Alzheimer’la nedensel ilişkisi bulundu. (Sochocka et al., 2017), Alzheimer’la nedensel ilişkili patojenleri şöyle listeliyor.
(Ledgerwood et al., 2003), “Yeni Virüs Teorisi” varsayımlarından yola çıkarak şizofreninin de patojenlerle ilişkili olabileceğini ileri sürüp aday olarak Toxoplasma Gondii’yi öneriyor. T. Gondii, bulaştığı memelilerde davranış ve beyin yapısı değişikliğine yol açabiliyor ve kedilerde sıkça görülüyor. Alakalı olarak, kedi sahipliğinin yaygın olduğu bölgelerde şizofreni de benzer biçimde yaygın. Dahası, şizofreni ve T. Gondii ilişkisine dair bir meta-analiz çalışması (Torrey et al., 2007), şizofreni hastalarında T. Gondii antikorlarının, kontrol gruplarına göre daha yüksek olduğunu (2.73 OR=) ve bunun şizofreniyle ilişkili bilinen tüm etkenlerden daha yüksek bir ilişkide olduğunu söylüyor. Henüz T. Gondii ve şizofreni ilişkisi kesin olarak kanıtlanamasa da bu, aralarında bir bağ olduğuna işaret ediyor.

Bunlar dışında da pek çok rahatsızlıkla patojenleri ilişkilendirebiliyoruz ve ilişkilendirmeye de devam edeceğiz gibi duruyor.
Peki, eşcinselliğin de büyük ölçüde patojen kaynaklı olmasını beklemeli miyiz?
Yaygın mı? Evet.
Evrimsel maliyeti var mı? Epey yüksek.
Nedeni hakkında ciddi bir fikrimiz var mı? Hayır. (Cinsel yönelimler ve nedenleri ile ilgili güncel inceleme makalesi için bkz: (Bailey et al., 2016; Bogaert ve Skortka, 2020))
Eşcinsellik, teorik olarak patojen teorisiyle en uyumlu kondisyonlardan. Dolaylı bir delil olarak, doğada eşcinselliğin görüldüğü sadece iki türden biri olmamız ve diğer türün de bizim evcilleştirdiğimiz koyunlar olması gösterilebilir. (Roselli ve Shormsak, 2009), tıpkı insanlardaki gibi koçlarda da eşcinselliğin ve aseksüelliğin yaygın olduğunu gösteriyor.
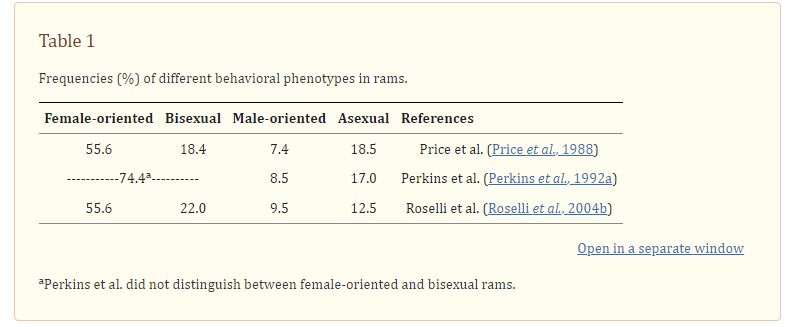
İnsanlar ve koyunlar arasında patojen geçişkenliği nadir olmadığı için bu patojen teori lehine yazılabilir. Eğer bu anormal davranışı koçlarda da görmeseydik sadece insan türüne yönelik açıklamaların (örneğin epigenetik veya doğumöncesi hormonal açıklamalar) gücü, patojen teorisine nazaran daha yüksek olabilirdi; fakat yakın ilişkide bulunduğumuz başka bir hayvanda da görünmesi ortak bir etiolojiye işaret ediyor gibi.
Eğer insanlarda eşcinsellik vakalarının çoğunluğu patojen kaynaklıysa, bunun ya doğum öncesinde ya da doğumdan hemen sonra bulaştığını söyleyebiliriz, zira eşcinsel erkeklerin beyni daha feminize ve bu feminizasyon daha çocukken bile kendini gösteriyor. Biraz daha spekülasyonda bulunursak, erişkin t-hücreli lösemi benzeri bir mekanizmanın işlediğini tahmin edebiliriz. Japonya’da sıkça çalışılan THLB, bebeklere anne sütünden geçiyor ve onlarca yıl boyunca (hastaların yarısından çoğunda 60 yaşa kadar) atıl halde bekleyip, sonra harekete geçebiliyor. Benzer bir şekilde, doğum öncesinde veya hemen sonrasında bulaşan patojenin beyin yapısını değiştirmesi mümkün.
Fakat bilim insanları bu konu üzerine eğilip patojenlerin eşcinsellik üzerinde rol oynayıp oynamadığını araştırana kadar bunların hepsi teorik spekülasyonlardan ibaret. Zayıf spekülasyon değil ama yine de spekülasyon.
Kaynakça
Apostolou, M. (2022). The Direct Reproductive Cost of Same-Sex Attraction: Evidence from Two Nationally Representative U.S. Samples. Archives of Sexual Behavior, 51(4), 1857-1864. https://doi.org/10.1007/s10508-021-02199-y
Bailey, J. M. (2009). What is Sexual Orientation and Do Women Have One? Içinde D. A. Hope (Ed.), Contemporary Perspectives on Lesbian, Gay, and Bisexual Identities (C. 54, ss. 43-63). Springer New York. https://doi.org/10.1007/978-0-387-09556-1_3
Bailey, J. M., Vasey, P. L., Diamond, L. M., Breedlove, S. M., Vilain, E., & Epprecht, M. (2016). Sexual Orientation, Controversy, and Science. Psychological Science in the Public Interest, 17(2), 45-101. https://doi.org/10.1177/1529100616637616
Bailey, J. V. (2003). Sexual behaviour of lesbians and bisexual women. Sexually Transmitted Infections, 79(2), 147-150. https://doi.org/10.1136/sti.79.2.147
Bjornevik, K., Cortese, M., Healy, B. C., Kuhle, J., Mina, M. J., Leng, Y., Elledge, S. J., Niebuhr, D. W., Scher, A. I., Munger, K. L., & Ascherio, A. (2022). Longitudinal analysis reveals high prevalence of Epstein-Barr virus associated with multiple sclerosis. Science, 375(6578), 296-301. https://doi.org/10.1126/science.abj8222
Bobrow, D., & Bailey, J. M. (2001). Is male homosexuality maintained via kin selection? Evolution and Human Behavior, 22(5), 361-368. https://doi.org/10.1016/S1090-5138(01)00074-5
Bogaert, A. F., & Skorska, M. N. (2020). A short review of biological research on the development of sexual orientation. Hormones and Behavior, 119, 104659. https://doi.org/10.1016/j.yhbeh.2019.104659
Cochran, G. M., Ewald, P. W., & Cochran, K. D. (2000). Infectious Causation of Disease: An Evolutionary Perspective. Perspectives in Biology and Medicine, 43(3), 406-448. https://doi.org/10.1353/pbm.2000.0016
Diamant, A. L., Schuster, M. A., McGuigan, K., & Lever, J. (1999). Lesbians’ Sexual History With Men: Implications for Taking a Sexual History. Archives of Internal Medicine, 159(22), 2730. https://doi.org/10.1001/archinte.159.22.2730
Ewald, P. W. (2002). Plague time: The new germ theory of diease. Anchor Books.
Ewald, P. W. (2009). Chapter 2 An Evolutionary Perspective on Parasitism as a Cause of Cancer. Içinde Advances in Parasitology (C. 68, ss. 21-43). Elsevier. https://doi.org/10.1016/S0065-308X(08)00602-7
Ganna, A., Verweij, K. J. H., Nivard, M. G., Maier, R., Wedow, R., Busch, A. S., Abdellaoui, A., Guo, S., Sathirapongsasuti, J. F., 23andMe Research Team, Lichtenstein, P., Lundström, S., Långström, N., Auton, A., Harris, K. M., Beecham, G. W., Martin, E. R., Sanders, A. R., Perry, J. R. B., … Zietsch, B. P. (2019). Large-scale GWAS reveals insights into the genetic architecture of same-sex sexual behavior. Science, 365(6456), eaat7693. https://doi.org/10.1126/science.aat7693
Ledgerwood, L. G., Ewald, P. W., & Cochran, G. M. (2003). Genes, Germs, and Schizophrenia: An Evolutionary Perspective. Perspectives in Biology and Medicine, 46(3), 317-348. https://doi.org/10.1353/pbm.2003.0038
Moran, P. A. P. (1972). Familial Effects in Schizophrenia and Homosexuality. Australian & New Zealand Journal of Psychiatry, 6(2), 116-119. https://doi.org/10.3109/00048677209159690
Roselli, C. E., & Stormshak, F. (2009). The neurobiology of sexual partner preferences in rams. Hormones and Behavior, 55(5), 611-620. https://doi.org/10.1016/j.yhbeh.2009.03.013
Sochocka, M., Zwolińska, K., & Leszek, J. (2017). The Infectious Etiology of Alzheimer’s Disease. Current Neuropharmacology, 15(7). https://doi.org/10.2174/1570159X15666170313122937
Sylva, D., Safron, A., Rosenthal, A. M., Reber, P. J., Parrish, T. B., & Bailey, J. M. (2013). Neural correlates of sexual arousal in heterosexual and homosexual women and men. Hormones and Behavior, 64(4), 673-684. https://doi.org/10.1016/j.yhbeh.2013.08.003
Torrey, E. F., Bartko, J. J., Lun, Z.-R., & Yolken, R. H. (2007). Antibodies to Toxoplasma gondii in Patients With Schizophrenia: A Meta-Analysis. Schizophrenia Bulletin, 33(3), 729-736. https://doi.org/10.1093/schbul/sbl050
Vasey, P. L., Parker, J. L., & VanderLaan, D. P. (2014). Comparative Reproductive Output of Androphilic and Gynephilic Males in Samoa. Archives of Sexual Behavior, 43(2), 363-367. https://doi.org/10.1007/s10508-013-0195-9
Whitehead, N. E. (2011). Neither genes nor choice: Same-sex attraction is
mostly a unique reaction to environmental factors. Journal of Human
Sexuality, 3, 81–114.